近日,植被结构功能与建造全国重点实验室/古生物研究院冯卓研究员应国际著名学术期刊Current Biology(《当代生物学》)邀请,为“Plant Evolution(植物演化)”专辑撰写综述导读Mass extinctions and land plant evolution(《生物大灭绝与陆生植物演化》)。在该文中,冯卓研究员结合自身多年研究积累,介绍了地质历史时期的生物灭绝事件以及陆生植物的响应,重点聚焦二叠纪末大灭绝事件对陆生植物演化造成的深远影响,并以化石记录最完整、研究最深入的陆地生态系统之一——华夏植物区为例,详细揭示了重大地质事件发生后陆地植被从崩溃到重建的全过程。
研究表明,几乎所有现代动物门类都在约5.18亿年前的寒武纪早期突然出现,这一事件被称为“寒武纪大爆发”。在此里程碑式的生物演化事件之后,海洋无脊椎动物的化石记录中出现了五次重大生物灭绝事件,合称“五大灭绝事件”,分别为奥陶纪末、晚泥盆世、二叠纪末、三叠纪末和白垩纪末的大灭绝事件(图1)。其中,二叠纪末大灭绝事件最为严重,在极短的地质时间内造成了约81%的海洋物种消失。
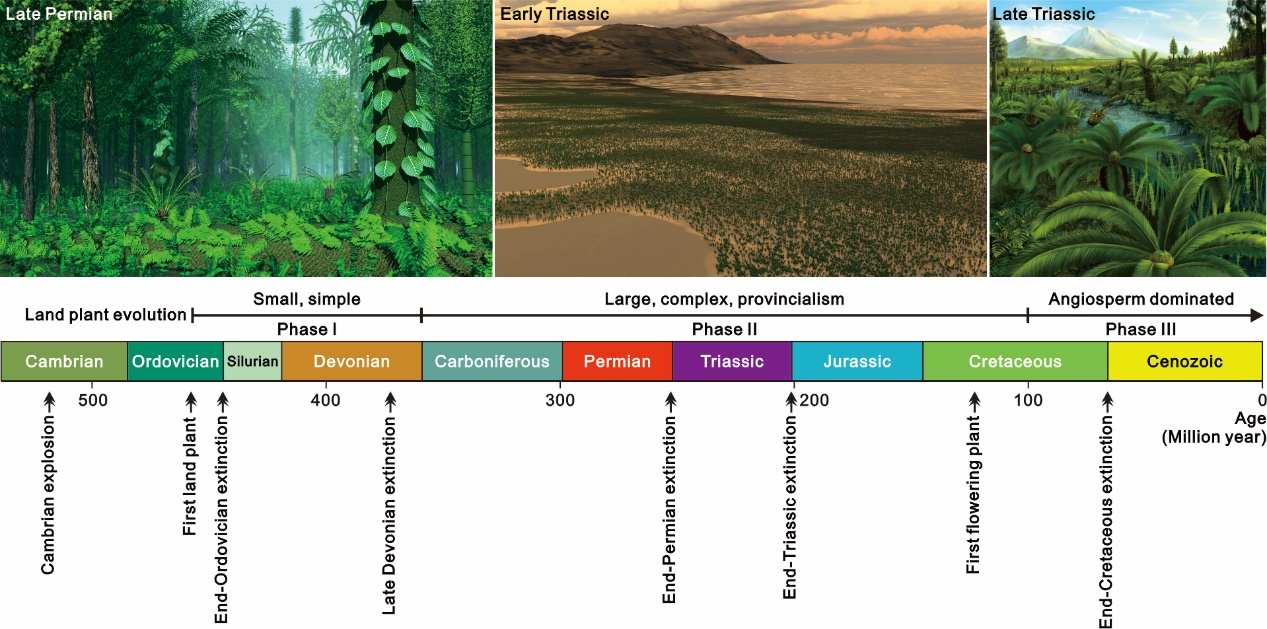
图1.重大生物演化事件与植物阶段性演化。
关于五大灭绝事件的根本诱因,目前尚无定论。然而,一个共同的显著特征是,这些事件都伴随突发性的全球环境与气候变化,且多数与大规模火山活动密切相关。奥陶纪末大灭绝发生时,位于南半球高纬度地区的冈瓦纳大陆经历了强烈的火山活动与冰川作用,随后引发的全球性降温导致海平面下降约100米。晚泥盆世大灭绝同样与可能起源于西伯利亚的大规模火山活动有关,之后地球再次经历了显著的降温事件。二叠纪末大灭绝是地球历史上最严重的一次灭绝事件,期间西伯利亚大规模火山喷发向大气中释放了大量有毒气体,并将大气CO₂含量推高至约2000–7000 ppm。三叠纪末大灭绝亦伴随着中大西洋岩浆省的大规模火山活动。白垩纪末大灭绝则由一颗小行星撞击地球引发,其证据是位于墨西哥湾、直径约200–300千米的希克苏鲁伯撞击坑。这一撞击事件可能进一步加剧了火山活动,并形成了类似“核冬天”的环境效应,最终导致非鸟类恐龙灭绝。
目前,陆生植物为何对每次海洋生物大灭绝事件表现出截然不同的响应,尚不明确。奥陶纪末大灭绝期间,植物刚登陆不久,仅是陆地生态系统中的次要组成部分,因此不易受突发性环境与气候变化的冲击。化石记录显示,晚泥盆世大灭绝并未引起陆生植物的衰退,反而在一定程度上促进了其演化;这一时期,小型树状石松类植物已形成局部的类森林生态系统,同时种子植物也开始多样化发展。被子植物(开花植物)在白垩纪末大灭绝发生期间仍保持高速辐射演化,最终成为陆地生态系统中的主导类群。然而,陆生植物在二叠纪末大灭绝期间表现出的灾难性衰退,与其他大灭绝事件中的情形形成鲜明对比,至今仍是一个未解之谜。一种较为合理的解释是,该事件中环境扰动发生得过于迅猛且程度空前严重,超出了陆生植物的适应极限。
陆生植物的阶段性演化。化石证据显示,陆生植物最早出现于奥陶纪中晚期(约460–470 Ma),标志着地球陆地植被化进程的开端。从宏观尺度来看,依据结构、功能及生态特征,陆生植物的演化大致可划分为三个主要阶段:前石炭纪、石炭纪—早白垩世以及白垩纪中期至今(图1)。石炭纪以前的陆生植物体型较小、结构简单,以石松类以及蕨类和种子植物的祖先类型为主。进入石炭纪,陆生植物发巨型化,高大的乔木型石松类(可高达50 m)和木贼类植物(可达30 m)与分异度极高的蕨类、种子蕨及原始裸子植物共同形成了广阔而复杂的森林生态系统,这一阶段的植物构成了地球上主要煤炭资源的物质来源。该时期,在气候和地理因素的共同作用下,形成了显著的植物地理区系格局,包括特提斯洋西侧中低纬度地区的欧美植物区、特提斯洋东侧中低纬度地区的华夏植物区、南半球高纬度地区的冈瓦纳植物区,以及北半球高纬度地区的安加拉植物区。自白垩纪中期以来,被子植物迅速辐射,最终成为全球植被的主导类群。
五大灭绝事件显著影响了海洋动物及海洋生态系统的演化轨迹,但似乎只有二叠纪末大灭绝引发了陆生植物群落的灾难性崩溃。晚古生代四大植物地理区系如何响应二叠纪末生物大灭绝事件,目前尚不清楚。由于二叠纪—三叠纪界线附近含植物化石的地层较为缺乏,且这些沉积地层尚缺乏高精度年代测定,因此,欧美植物群和安加拉植物群的灭绝时间与过程至今仍存在争议。相比之下,冈瓦纳植物群和华夏植物群的研究程度较高,其年代框架也得到较好的限定。尤其华夏植物区,保存了唯一连续跨越早三叠世至晚三叠世的植物化石记录,为二叠纪末大灭绝之后森林生态系统的逐步恢复、辐射演化以及重新建立提供了重要实证。
华夏植物群。自石炭纪中期出现以来,华夏植物群便以独特的东方型植物类群为重要特色,包括乔木型石松类、木贼类、前裸子植物、蕨类、苏铁类以及高度多样化的大羽羊齿类植物(图2),整体呈现为热带至亚热带植被类型。其中,大羽羊齿类最具代表性,其最显著特征是具有类似被子植物的大型复叶,并拥有复杂的网状脉序,最高可达四级脉序(图3)。由于大羽羊齿类在华夏植物群中占据优势且广泛出现,华夏植物群亦常被称为“大羽羊齿”植物群。化石证据表明,部分大羽羊齿类植物具有钩状附属结构,暗示其可能具有攀援性或藤本生长习性。基于一种特殊昆虫取食损伤结构的新研究成果表明,部分大羽羊齿类植物具有感夜性(睡眠)运动,这一较为先进的生态生理表明,该类适应特征在不同植物谱系中或是独立演化而来,属于典型的趋同演化结果。

图2(左).华夏植物群的代表性植物。(A) Lepidodendron oculus-felis. (B) Lobatannularia sp. (C) Pecopteris sp. (D) Tingia carbonica. (E) Gigantonoclea guizhouensis.
图3(右).大羽羊齿类叶片多级网状脉序(A)和分泌结构(B)。
华夏植物群广泛分布于今天的东亚(中国、朝鲜、韩国、日本)、东南亚(越南、老挝、泰国、马来西亚、印度尼西亚和新几内亚)以及中东地区(伊拉克、约旦、沙特阿拉伯、阿曼和土耳其)。其中,中国保存了华夏植物群最完整的演化序列记录,涵盖了从起源、经演化鼎盛到最终灭绝的全过程。此外,华南板块三叠纪地层还保存了一系列植物化石组合,为揭示陆生植物对二叠纪末灭绝事件的响应,以及其后植被恢复、辐射演化和森林生态系统重建的阶段性过程,提供了独特契机。高精度锆石U-Pb测年显示,华夏植物群的最后出现时间约为251.7 Ma,即比二叠纪—三叠纪界线年轻约20万年,表明华夏植物群在消亡之前一直延续到了三叠纪初期。值得注意的是,与之相比,冈瓦纳地区的舌羊齿植物群的灭绝发生在二叠纪—三叠纪界线之前约30万年,与海洋动物群灭绝事件的时间一致。低纬与高纬陆地生态系统对二叠纪末大灭绝表现出这种时间差异的原因仍不清楚,但部分学者推测,一系列短暂的温度剧烈升降事件可能是造成这一差异的重要驱动因素。
华夏植物群的消亡。至二叠纪末期,原本广泛分布的华夏植物群仅残存于华南板块的西南缘,即今天的中国西南地区。在该地区,以大羽羊齿类、乔木状石松类、木贼类、蕨类、种子蕨以及少量原始裸子植物为代表的植物类群,构成了局部分布的类雨林生态系统。然而,到二叠纪—三叠纪之交,在高地环境中,多样化的植被群落首先被以盾籽类种子蕨占优势的灌丛植被所替代;随后,在早三叠世早期(印度期早期),华夏植物群在低地环境中最终被由草本石松类组成的单一种群彻底取代,标志着华夏植物群的最终消亡。在不同地质剖面中,含草本石松类植物化石的层位均位于华夏植物群最后出现层位之上约1.5–2 米处,表明在华夏植物群灭绝之后的很短时间内植被便开始重新恢复了。
早三叠世植被的辐射演化。在含草本石松类(卡以头组)之上的东川组,保存了一个以松柏类植物为主的植物组合,时代属早三叠世早期(印度期晚期)。该组合多样性低,除了丰富的原始松柏类(Albertia)外,还包含少量石松类(Lepacyclotes型)、木贼目(Paracalamites)、紫萁目(Anomopteris)等(图4)。在华南板块下三叠统上部(奥伦尼克阶),还发育了一个组成上相似但多样性显著更高的植物组合,包括木贼类、蕨类、松柏类和苏铁类植物,共28属38种。就丰度和多样性而言,松柏类植物,尤其是Albertia和Voltzia,是最重要的组成分子。华南板块的奥伦尼克期植物群,在组成上与欧洲同期的斑砂岩植物群相似。

图4.早—中三叠世代表性植物。(A,B) Albertia spp. (C) Anomopteris sp. (D) Paracalamites sp. (E) Tomiostrobus sinensis. (F) Lepacyclotes sp.
中三叠世的过渡型植被。中三叠世植物多样性总体上与奥伦尼克期植物组合相近,但其组成面貌却发生了显著变化。华南板块中三叠世植物化石组合以多样的石松类植物Pleuromeia和Lepacyclotes为主,它们是绝对优势类群。在中三叠世,这两类石松类植物广泛分布于整个北半球,通常被视为环境胁迫耐受力强的植物类群。华南板块中三叠世植物组合中还包含多样性较低的木贼类、盾籽类种子蕨、松柏类和苏铁类植物。中三叠世植物组合与奥伦尼克期植物组合在组成面貌上的差异表明,早三叠世的植物辐射演化可能只是植物演化史中一个短暂且未能延续的发展阶段。
晚三叠世森林生态系统的重建。华南板块广泛发育一类高度多样化的晚三叠世植物组合,通常称为Dictyophyllum–Clathropteris植物组合。晚三叠世植被的种类丰富度已可与晚二叠世华夏植物群相媲美,但以分异度极高的双扇蕨科(即Dictyophyllum 和Clathropteris)、苏铁目和本内苏铁目植物为重要特色,其中后两类群在某些化石组合中可占总植物多样性的50%以上。苏铁目和本内苏铁目在快速辐射演化之后,最终成为中生代植被中的优势类群。因此中生代常被称为“苏铁时代”。
晚三叠世植物组合已呈现出较为明显的现代面貌,其中包含若干现代类群,尽管其多样性仍较低;这些植物包括木贼类、若干蕨类谱系、原始银杏类以及松柏类植物(图5)。曾经繁盛于广布而复杂的森林生态系统中的晚三叠世植物群,为华南板块丰富的煤炭资源的形成作出了重要贡献。有趣的是,华南板块的化石记录证实,双扇蕨科植物被昆虫留脉式取食的最早证据可追溯至晚三叠世;同时,一些高度复杂的昆虫繁殖策略(如植内产卵)和取食策略(如对虫卵的刺吸式取食行为)都在这一时期出现。值得注意的是,最新研究表明,晚三叠世的银杏类植物已具有针对昆虫潜叶取食的化学防御能力。这些发现表明,晚三叠世陆地生态系统已经达到相当高的复杂化程度。

图5.晚三叠世代表性植物。(A) Dictyophyllum nathorstii (Dipteridaceae). (B) Clathropteris meniscioides (Dipteridaceae). (C) Coniopteris sp. (Dicksoniaceae). (D) Cladophlebis raciborskii (Osmundales). (E) Baiera multipartita (Ginkgoaceae). (F) Pterophyllum schenkii (Bennettitales)
结语。华夏植物群灭绝之后,植被的逐步恢复与演替过程,在华南板块得到了独一无二的记录,为我们理解陆地生态系统在灾难性崩溃后如何实现重建提供了关键范式。这一过程揭示了灭绝的选择性、幸存者适应性,及其在生态复杂性恢复所需漫长时间尺度之间错综复杂的关系,也为当前气候变化与生物多样性挑战提供了重要启示。
关于人类是否正步入第六次生物大灭绝事件,学界目前仍存在争议。然而,毋庸置疑的是,我们正在目睹全球生物多样性的急剧丧失——这或许是人类所面临的最严峻挑战之一。与由自然过程驱动的史前五大灭绝事件不同,当前的生物多样性丧失主要是由人类活动造成,如土地利用变更和森林砍伐。每个物种都与无数其他物种相互作用,共同编织成复杂的生态系统网络,而这一系统反过来又维持折其中所有生命的生存需求。因此,即使单个物种的灭绝或其种群数量的显著下降,也可能波及其他物种,进而削弱整个生态系统的功能及其所能提供的资源。从这个意义上说,保护全球生物多样性,本质上就是保护人类自己。
论文信息:Feng, Z., 2026. Mass extinctions and land plant evolution. Current Biology 36, R491–R521. https://doi.org/10.1016/j.cub.2026.01.037
来源:古生物研究院
编辑:张懿淼 责任编辑:李哲

 En
En